중추신경계통 의 통합기능
신경생리학 : 함기선. 신문균. 최흥식 공저, 현문사, 1997, Page 237~268
|
1. 대뇌피질의 구조와 일반 생리학 : 뇌파 (뇌전도) 4) 뇌파 (뇌전도) Electroencephalogram (EEG) |
중추신경계통의 통합기능에는 감각입력의 처리기구, 운동 및 자율활동의 처리기구 등의 분류에 직접 해당하지 않는 중요한 과정이 포함되어 있다. 이와 같은 신경기구는 학습, 수면과 각성 주기 cycle, 의식 언어 및 기억 등의 기초로 되어 있다. 그 외에 원시적 또는 기본적 행동 형태 elementary behavior pattern 및 정서의 기초로 되어 있는 통합기구가 있다. 하지만 다른 통합기구, 예를 들면 복잡한 행동형태의 기초를 이루는 신경생리학적 기구 등에 대해서는 현재 불분명한 점이 많다.
중추신경계통의 통합기구에 관여하는 중요한 부분은 대뇌피질 cerebral cortex 이다. 대뇌피질은 계통발생학적으로 가장 새로운 것이고 사람에서는 매우 잘 발달되었으며 이 때문에 많은 주름이 겹쳐 있는 모양을 하고 두개골 속에 들어 있다.
중추신경계통의 통합작용에 있어서 감각계통 sensory system 은 운동계통이나 자율신경계통이 똑같이 불가결한 것이다. 감각계통이란 신체내외의 정보를 받는 수용기, 정보를 전달하는 구심성 신경섬유, 정보를 처리하는 감각중추의 3 과정으로 이루어지는 말초성 및 중추성의 신경계통기구인 것이다. 운동계통 및 자율신경계통에 대해서는 지금까지 상세히 설명했고 감각계통에 대해서는 제 6 장과 제 7 장에서 설명했다. 본 장에서는 중추신경계통의 통합기능과 감각계통의 관련에 대해 언급하지만 필요에 따라 제 6 장을 참고해 주기 바란다.
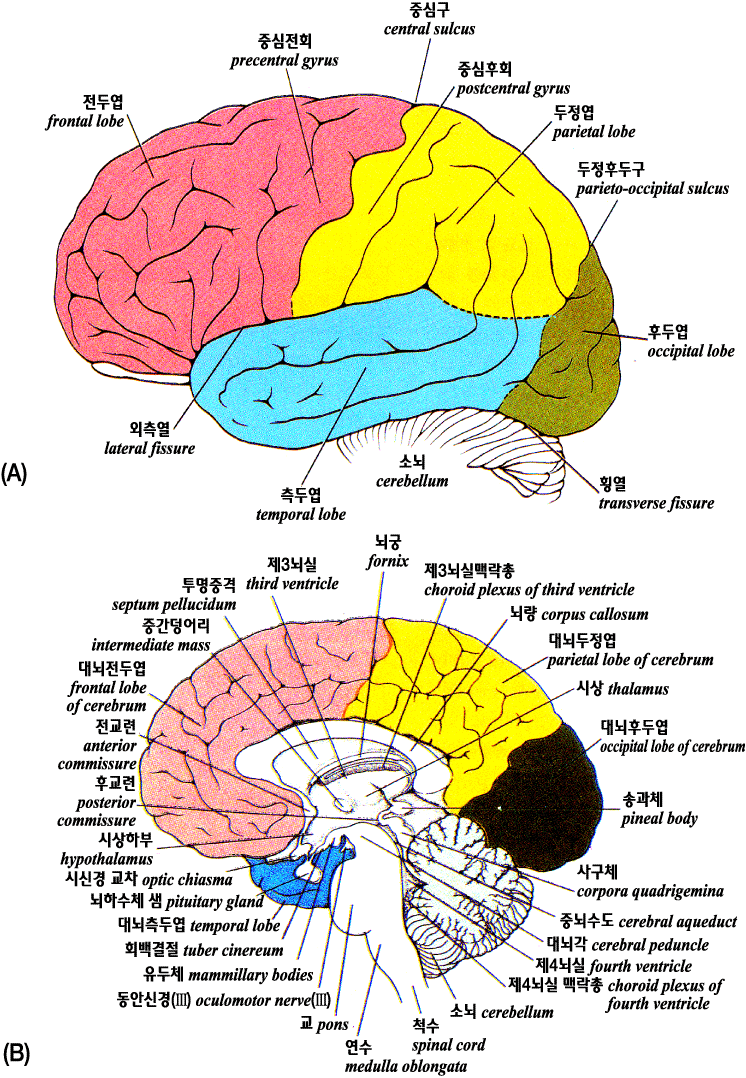
사람 대뇌의 좌우 반구 hemisphere 는 두 개강의 대부분을 차지하고 있다. 대뇌피질의 표면은 구 sulcus 에 의해 서로 떨어져 있는 회 gyrus 라고 하는 볼록면의 집합으로 되어 있고 복잡한 주름형태를 하고 있다. 다음 그림에 나타난 것같이 뇌간과 간뇌(그 중 중요한 부분은 시상과 시상하부이다) 는 대뇌피질에 의해 둘러싸여 있다. 이들의 상대적인 위치는 정중시상단면, 수평단면 및 전두면에서 이해될 수 있다.
대 뇌
A:
외측면, B: 내 면
좌우 대뇌반구는 4 개의 엽 lobe 으로 크게 구분된다. 두개 내 에서의 위치로부터 4 개의 엽을 각각 전두엽, 두정엽, 측두엽, 및 후두엽이라고 한다. 각각의 엽은 몇 개의 회로 구성된다. 엽의 분류는 전적으로 해부학적인 것으로 각 엽의 기능을 말하는 것은 아니다.
대뇌피질은 각 영역의 구심성 및 원심성 섬유의 연락에 의해 기능적으로 분류할 수 있다. 이와 같은 분류는 단순하지만 유용하고 뇌손상에 따르는 임상적 탈락증상의 관찰로부터 얻을 수 있는 소견과도 일치한다. 예를 들면 중심전회와 그 주변부는 특수 운동피질 specific motor cortex 이고 여기에서 나오는 원심성 신경섬유가 내포되어 한쪽만 절단되면 경련성 편마비라고 하는 심한 운동기능의 장애가 일어난다.
시상감각핵을 경유한 후에 감각로가 끝나는 피질영역을 특수 감각피질 specific sensory cortex 라고 한다. 이들 특수영역은 사람을 포함해 4 종류의 동물의 뇌반구 위에 나타난다. 예를 들면 두정엽의 중심후회는 체성 감각로가 끝나는 가장 중요한 영역이다. 후두엽에는 시각로, 측두엽에는 청각로가 끝나므로 각 영역을 시각피질, 청각피질이라고 한다. 이들의 특수 감각 피질영역의 손상은 그것이 관장하는 특정감각의 장애를 일으킨다. 예를 들면 중증 예에서는 피질맹 cortical blindness, 피질성 난청 cortical deafness 이라고 하는 중추성의 장님이나 귀머거리로 된다.
대뇌피질의 모든 부분이 확인되고 있는 전도로의 주행에 기초를 두어 특수감각영역과 운동영역으로 분류되는 것은 아니다. 그 어느 것에도 속하지 않는 부분이 있고 그 부분은 주로 운동영역과 감각영역의 상호작용에 관여한다고 생각되고 보통 연합피질 association cortex, 때로는 비특수피질 non-specific cortex 이라고도 한다.
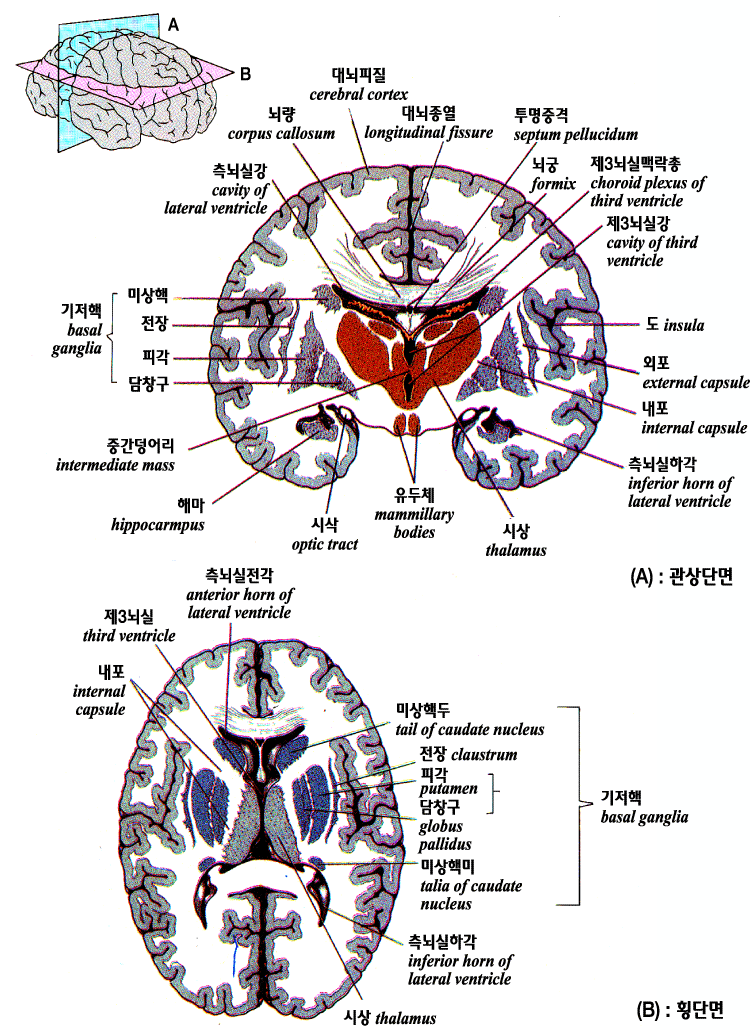
뇌의 관상단면과 횡단면
연합피질은 사람에서는 물론 유인원에 있어서도 위의 그림에 나타난 것처럼 감각 및 운동피질에 비해 광범위한 부분을 차지한다. 더구나 뇌의 크기는 진화과정에서 증대해 가고 있다. 이들 2 개의 요소에 의해 사람의 연합피질의 절대적 및 상대적인 크기는 증대되어 있다. 연합피질은 앞에서 정의한 중추신경계통의 고도의 통합기능에 중대한 역할을 하고 있다.
대뇌피질은 피질하부로부터 오는 구심성 섬유를 대부분 시상피질로 thalamocortical tract 를 개재해서 받는다. 모든 신경계통 및 감각기로부터 대뇌로 가는 입력은 최종적으로 시상에서 신경원을 중계한 후 피질에 투사된다. 피질에서 오는 원심성 섬유를 투사섬유 projection fiber 라고 한다. 연합섬유 association fiber 와 교련섬유 commissural fiber 는 특히 중요하다. 연합섬유는 한쪽의 반구내에서 피질과 피질을 연락하고 있고 교련섬유는 주로 뇌량을 경유해서 양쪽의 반구피질을 연락하고 있다. 연합섬유와 교련섬유는 피질영역의 어느 것인가를 중심으로 하고 있다고 생각되명 구심성 또는 원심성으로 나눌 수 있다.
대뇌피질은 표면적이 약 2,200㎠ (47 × 47 ㎝) 의 얇은 층의 신경조직으로 구성되어 있다. 층의 두께는 장소에 따라 1.3 ㎜ 로부터 4.5 ㎜ 까지 변화가 많다. 대뇌피질의 부피는 약 600 ㎤ 이고 10억개 내지 100 억개의 신경세포 (신경원) 와 다수의 신경교세포 neuroglial cell 를 함유하고 있다. 신경교세포의 정확한 수는 알 수 없다. 피질내부에는 주로 세포체를 함유하고 잇는 층과 주로 축삭을 함유하고 있는 층이 서로 엇갈려 존재하고 있다. 이 때문에 신선한 피질의 단면은 줄무늬 모양을 나타내고 있다. 전형적인 대뇌피질은 세포의 형태 및 배열에 의해 6 층으로 식별되지만 이들의 구조에 대해서는 여기서는 자세히 설명하지 않는다.
조직학적 및 전기생리학적 관찰에 의하면 대뇌피질에서의 정보처리는 기본적으로 대뇌피질표면에 대해 수직으로 배열되어 있는 신경회로에 의해서 이루어진다. <그림 4B> 의 단순화한 모식도에서 그 배열을 나타냈다. 표면의Ⅰ층에서 Ⅳ층은 주로 피질로 입력되는 정보의 수리와 처리에 관여한다. 대뇌피질에서 오는 원심성 신경원의 세포체 (투사, 연합 및 교련섬유) 를 함유하고 있다. <그림 3> 은 깊은 층의 Ⅴ층 내지 Ⅵ층에 존재하는 경향이 있다. 그러므로 Ⅴ층과 Ⅵ층을 피질성 원심로의 기시부로 볼 수 있다.

<그림 3> 대뇌반구에 있어서 섬유연락
연합섬유는 대뇌반구의 다른 부위을 서로 연락한다. 교련섬유는 양쪽의 대뇌반구를 (주로 뇌량을 통해서)연락한다. 대뇌반구는 피질원심성(투사) 섬유를 개재해서 다른 신경계의 연락한다. 대뇌반구로 가는 구심성 섬유는 시상피질로를 통과한다.
위에서 설명한 대뇌피질의 배열기구에 대해서는 이미 제 8 장에서 중심전회에 체성 기능의 국재기구가 있다고 설명했지만 똑같은 기구가 특수감각영역에 있어서도 볼 수 있다. 중심후회, 즉 체성 감각피질에는 중심전회와 유사한 체성 기능의 국재성 배열이 보이고 또한 다른 감각피질에서도 똑같은 (때로는 복작한) 배열이 보인다. 이와 같이 어떤 기능에 관련된 신경원은 피질표면에 대해 수직으로 정렬해서 원기둥 모양으로 몰려 있다. 이러한 기능적으로 관련된 어느 세포군을 피질원주 cortical column 라고 한다. 운동피질의 미소부위 자극 및 기록의 실험에 의해 이들의 기능적 피질원주의 직경은 약 1 ㎜ 라는 것이 밝혀졌다. 그러나 피질신경원의 연결이나 투사에 대해서는 현재까지 거의 알려진 것이 없다.

<그림
4> 대뇌피질의 층구조의 개략도
A: 왼쪽-Golgi 염색표본, 개개의 신경원과
수상돌기가 강조되고 있다. 가운데-Nissl 표본, 세포체만이
염색되어 있다.
오른쪽-myelin 초 표본 축삭의 경로가 보인다.
B: 대뇌피질 표면에 대해 수직으로 배열된 신경원정보의
흐름 (적색) 의 모식도, 추체세포와 성상세포만을 나타냈다.
흥분성
연접을 + 로, 억제성 연접을 - 로 나타냈다.
1개의 전극을 두피 위에, 다른 전극 (불관전극 indifferent electrode) 을 귓바퀴에 붙이면 2 개 전극 사이의 전압차가 기록된다. 이러한 연속적인 전압기록을 뇌파 (뇌전도, electroencephalogram, EEG) 라고 한다. 뇌파는 1 ~ 50 ㎐ 의 주파수와 약 10 ~ 10 ㎶ 의 진폭을 갖는다. 뇌파는 독일의 신경생리학자 Hans Berger 에 의해 처음 시도되었다. 그는 1929 ~ 1938 년 사이에 뇌파를 임상적 또는 실험적으로 응용하기 위한 기초적 연구를 했다.
뇌파는 보통 신경학적 (그림 5 왼쪽) 와 기록의 조건 (예를 들면 기록지의 속도나 증폭기 계통에 있어서 filter의 주파수 응답)등은 포준화되어 왔다. 기록은 단극도출 unipolar lead 또는 쌍극도출 bipolar lead 법에 의해 이루어진다. 기록의 평가는 파형의 주파수, 진폭, 형태, 이상파형의 출현빈도 및 위치적 분포에 대하여 육안으로도 할 수 있는 것도 있고 analogue 또는 digital computer 로 해석할 수도 있다.
뇌파의 주파수와 진폭은 여러 인자로 결정된다. 기록부위는 <그림 5> 에 나타난 것같이 그 중 하나의 인자이다. 예를 들면 눈을 감았을 때 후두부에서 기록되는 뇌파의 진폭은 전두부나 두정부의 것보다 크다. 뇌파의 파형은 깨어있는 상태 wakefulness 의 정도에도 영향을 미친다 (그림 5).
눈을 감았을 때 진폭이 큰 저주파의 파형은 눈을 뜨는 동시에 진폭이 작은 고주파의 파형으로 바뀌고 눈을 감은 후에 다시 저주파의 파형이 시작된다. 이와 같은 저주파의 리듬은 건강한 성인이 각성 때에 눈을 감고 안정을 취한 상태에서 잘 볼 수 있다. 이것은 후두엽에서 가장명확하고 주파수 성분은 8 ~ 13 ㎐ (평균10㎐) 이다. 이와 같은 파형을 α 파는 <그림 5> 에 나타난 것과 같이 대부분의 기록 부위에서 거의 같은 파형 (진폭, 주파수, 위상) 으로 나타나며 이것을 동기화 뇌파 synchronized EEG 라고 한다.
α파는 눈을 떴을 때 (그림 5
의 중앙부의 기록) 나
다른 감각기관을 자극했을 때 또는 정신활동 때에는 소실된다. 이와 같은 소실을
α파차 단 alpha-blodking
이라고 한다. α 파차단 때에는 14
~ 30 ㎐ 의 높은 주파수 (평균 20 ㎐) 의 작은
진폭을 갖는  파 bet a wave
가 출현한다. 이 경우 뇌파의 파형은 불규칙하고 기록부위에 의해 진폭,
주파수, 위상이 크게 다르다. 이것을 뇌파의 탈동기 desynchronization
라고 한다.
파 bet a wave
가 출현한다. 이 경우 뇌파의 파형은 불규칙하고 기록부위에 의해 진폭,
주파수, 위상이 크게 다르다. 이것을 뇌파의 탈동기 desynchronization
라고 한다.
이상 설명한 뇌파의 파형 이외에도 다음 2
개의
중요한 파형이 있다. 그 하나는 4 ~ 7 ㎐ (평균 6 ㎐) 의 저주파로 큰 진폭을 갖는
 파 theta wave 이고, 다른 하나는 0.3
~ 3.5 ㎐ (평균 3 ㎐) 의 저주파로 큰 진폭을 갖는
파 theta wave 이고, 다른 하나는 0.3
~ 3.5 ㎐ (평균 3 ㎐) 의 저주파로 큰 진폭을 갖는 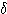 파 delta wave
이다.
파 delta wave
이다. 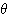 파와
파와 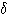 파는 정상 성인에서는 각성 때에 출현하지 않지만 수면시 볼 수 있다.
파는 정상 성인에서는 각성 때에 출현하지 않지만 수면시 볼 수 있다.
뇌파의 주파수와 진폭은 기록부위와 깨어 있는
상태의 요인 이외에도 동물의 종류나 나이에 따라서도 다르다. 예를 들면 사람의
유아기와 미성년기의 뇌파는 성인보다 저주파이며 한 편 불규칙하다. 어린
사람에서는 각성 중에도 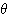 파나
파나 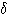 파를 기록할 수 있다.
파를 기록할 수 있다.
대뇌피질의 어떤 전기적 과정이 뇌파를 발생시키고 있는지 앞에서 설명했듯이 피질선경원에는 전도성 활동전압 및 국소성 느린 전압 slow potential 변동이 발생된다. 동물실험에 있어서 뇌파와 단일피질 신경원의 활동을 동시에 기록함으로써 이들 2 개의 전압과 전파의 관계가 연구되었다. 그 결과 뇌파는 명백히 피질신경원의 막전압의 느린 변화, 특히 흥분성 연접후전압 (EPSP) 과 억제성 연접후전압 (IPSP) 을 주로 반영하고 있다는 것을 알았다. 보통의 상태에서는 신경원 (또는 피질신경교세포) 내를 전도하는 활동전압에 의해서는 뇌파는 영향을 받지 않는다.
뇌파의 기록전극은 피질의 전류발생원 (EPSP, IPSP 발생 중에 막을 통과하는 이온전류) 으로부터 비교적 떨어져 있다. 따라서 뇌파의 진폭은 세포내 전극으로 기록되는 신경원 막전압 진폭의 1/100 ~ 1/1000이다 (그림 5, 9 와 그림 3-10, 11 의 종축에 나타난 것을 비교해 보시오).
뇌파를 동물실험이나 사람의 뇌의 외과수술중에 대뇌피질의 표면에서 직접 피질뇌파 electrocorticogram 로 기록하면 두피 위에서 기록 했을 때 보다도 10 배 정도 큰 진폭을 나타낸다. 어떤 경우에도 뇌파는 다수의 신경세포에 동시에 발생하는 전압변화를 반영하고 있다. 피질의 표면에 직접 접촉되어 있는 면적 1 ㎟의 전극은 깊이 약 0.5 ㎜ 까지의 약 100,000 개의 신경원 활동을 반영한다. 또한 기록이 두피 위에서 이루어질 때에느 그보다 약 10 배의 신경원이 뇌파의 진폭에 관여한다. 따라서 뇌파에 큰 진폭의 파형이 나타나기 위해서는 전극 밑의 다수의 신경원의 동시적 활동이 필요하다.
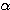 파 리듬과 같은 대뇌피질의 주기적 활동은 대뇌피질이 아니라 시상에서 유래한다.
리듬은 시상으로부터 피질로 전달된다. 시상피질로의 절단 또는
시상의 주기적 활동은 시상-피질연락의절단 혹은 피질제거 decrotication 후에도 남아 있고 시상으로 가는 입력에서 변화한다. 특히 뇌간망상체
brain stem reticular-forma-tion
는 동기화와 탈동기화와의 두 가지 작용을 가지며 시상의 주기적 전압활동을 증강시키거나
억제시키고 있다. 이에 대해서는 p.248 의 수면각성주기에는 자세히 설명한다.
파 리듬과 같은 대뇌피질의 주기적 활동은 대뇌피질이 아니라 시상에서 유래한다.
리듬은 시상으로부터 피질로 전달된다. 시상피질로의 절단 또는
시상의 주기적 활동은 시상-피질연락의절단 혹은 피질제거 decrotication 후에도 남아 있고 시상으로 가는 입력에서 변화한다. 특히 뇌간망상체
brain stem reticular-forma-tion
는 동기화와 탈동기화와의 두 가지 작용을 가지며 시상의 주기적 전압활동을 증강시키거나
억제시키고 있다. 이에 대해서는 p.248 의 수면각성주기에는 자세히 설명한다.
간질발작의 진단에 대표되는 뇌파의 임상적응용에 대해서는 본 장에서 취급하지 않았다. 뇌파의 소실 (뇌파가 평탄하게 전압을 나타낸다) 은 죽음의 지표 criterion of death 로서 일반적으로 받아들이고 있다. 진보된 소생술에 의해 순환부전 및 호흡부전으로부터 회복된 환자가 역시 의식회복과 자발호흡의 징후를 나타내지 않을 때 에는 허혈 (순환장애) 에 의해 대뇌피질 및 뇌간이 불가역적으로 장애될 가능성이 있다. 특히 어린 사람이 사고로 이와 같은 뇌사 brain death 에 빠질 경우 허혈이나 그에 의한 산소공급이 없는 상태에서도 보다 오래 기능을 유지할 수 있는 다른 기관 (예: 신장이나 심장) 은 적당한 조건이 조절되면 장기이식에 사용될 수 있다.

<그림 5> 각성 때의 안정상태에 있어서 사람의 정상 뇌파
위쪽의 그림에 표시한 두개골 위의 부위로부터 단극도출법에 의해 8 channe (통로) 을 동시에 기록, 전극의
1 개는 무관전극으로서 귀바퀴에 부착시킨다. 눈을 떴을 때에는 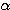 파가 중단된다
파가 중단된다
말초의 구심성 신경 (예: 피부신경, 그림 6) 또는 감각기관을 역치이상의 강도로 자극하면 이것에 대응하는 감각피질에 약 10 msec 의 잠복시간을 갖는 전압변화가 유발된다. 이 전압을 유발전압이라고 한다 (그림 6B, C). 최초로 양성으로 흔들리는 전압을 제 1 차 유발전압 primary evoked potential 이라 한다 (그림 6B). 이것은 피질의 한정된 좁은 부위, 즉 말초자극 부위에 대응하는 피질 투사영역 cortical projection area 에만 유발된다. 예를 들면 피부신경자극의 경우, 제 1 차 유발전압은 자극부위에 대응하는 중심후회의 체성 감각영역에만 유발된다. 제1차 유발전압 직후에 제2차 유발전압 secondary evoked potential 이라고 하는 약간 긴 반응이 유발된다 (그림 6C). 이와 같은 제2차 유발전압은 광범위한 대뇌피질로부터 기록된다.
유발전압의 기초를 이루는 신경기구에는 뇌파 파형의 경우와 똑같은 신경원의 흥분충동 impulse 활동은 아니고 오히려 느린 연접활동이 관여한다고 생각된다. 이 경우도 또한 전극이 기곡되고 있는 것은 다수 신경원의 세포외 전류에 의한 전압의 총화이다.
유발전압은 말초신경 또는 수용기의 자극 후 뿐만 아니라 중추의 전도로 또는 핵, 피질 등의 자극에 의해서도 유발된다. 유발전압은 말초와 중추의 연락 또는 중추의 다른 부위 상호연락을 연구하기 위한 전기생리학적 수법으로서 중요하다. 반복자극에 대한 반응을 가산 평균하는 방법 (이 과정은 computer 로 한다) 에 의해 뇌파 활동 파형에 묻혀 있는 약한 유발전압을 밝혀낼 수 있다. 가산평균방법을 응용한 정교한 실례로는 수의운동에 관련된 유발준비전압 evoked readiness potential 이다. 가산평균된 유발전압은 임상적으로도 응용될 수 있다. 예를 들면 음성자극에 대한 청각피질의 유발전압의 기록은 어린이들에게 있는 어떤 종류의 난청진단에 사용된다. 유발전압의 검사는 난청의 종류나 진행을 아는 것 외에 객관적 진단에도 쓰인다.

<그림
6> 사람의 뇌파로부터 유발, 기록된 유발전압
A: 실험배치도, 자극은 피부의
전기자극이다. 그러나 다른 자극 (예: 기계적 자극 또는 온도 자극) 도
사용할 수 있다.
전압은 두피 위의 뇌파용 전극에 의해 기록된다.
B: 중심후회의
대응하는 투사영역으로부터 기록된 제 1 차 유발전압
C: 제 1 차 유발전압 및 제2차
유발전압, B 와 C 에서는 시간눈금이 다른 것에 주의할 것
사람은 안정 때에는 1 분 당 전신에서 약 250 ㎖ 의 산소를 소비한다. 뇌의 산소소비량은 그 무게에 비해 크고 전신의 산소소비량의 약 20 % 를 차지한다. 즉 약 50 ㎖/분 의 산소가 뇌의 신경원과 신경교의 대사에 사용된다. 뇌 중에서도 대뇌피질의 산소소비량은 가장 많아 100 g 조직 당 약 1 ㎖/분 이다. 대뇌피질의 산소필요량이 가장 높은 것은 거의 8 ~ 12 초 동안 뇌로 가는 혈류가 정지되어 산소수송이 중단되면 의식소실이 일어난다고 하는 사실에도 반영되고 있다.
이와같이 산소소비량이 높게 유지되고 있는
것은 피질의 기능과 관계가 있다. 즉 피질의 어느부분의 활동이 증가하면 바로
그 부위의 국소산소소비량은 증대되고 동시에 국소혈류량이 중가하는 기구로 되어
있다. 혈류량의 증가는 <그림 7A> 에 나타난 방법 (D.H. Iogvar와 N.A.
Lassen 에 의해 개발되었다.) 에 의해 측정할 수 있다. 우선 방사선 키세논 가스
(¹³³Xe) 를
소량 경정맥으로 주입한다 (이것은 전혀 해가 없다). 측두부에 Geiger counter
(실제로
254 개도) 를 설치해서 이것에 의해 뇌의 각부 위에 도달한 키세논 가스를 검출한다.
이와 같은 방사선 강도는 직접 국소혈류량을 산출할 수 있다.
건강한 피험자에게 이 방법을 써서 측정한
결과를 <그림 7B> 에 요약해서 나타냈다. 뇌파가 전형적 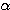 파를 나타내고 있는 안정시의 사람에게는 뇌의 전두부의 혈류량이 다른 부분보다도
현저히 놓다. 반대쪽 (이 경우는 오른쪽) 손의 피부에 통증을 동반하지 않는
자극을 주어도 뇌의 혈류분포의 형태는 거의 변화지 않는다 (그림 7B 의 촉각자극). 즉 통각을 동반하는 자극 (그림 7B
의
통각자극) 에서는 혈류의 분포는 기본적으로 변하지 않지만 뇌의 총 혈류랑(그림 7B
의
각 뇌의 위에 % 로 나타냈다) 은 명백히 증가한다. 반대쪽 손을 자발적으로 열었다 닫았다
하면 (손의 운동) 총 혈류량은 증가한다. 이 경우에는 중심후회의 체성 감각영역과
그 부근의 두정피질의 혈류량이 동시에 증대된다. 이야기를 하거나 독서를
하면 혈류량의 증대부는 I 형의 분포로 되고 특히 독서 때에는 혈류의 중대부가 후두엽의
시각영역에까지 확대된다. 생각을 하거나 수를 세거나 하는 실험에서는 뇌의
총혈류량이 증가하고 중심구의 전 및 후부의 혈류량이 증대하게 된다.
파를 나타내고 있는 안정시의 사람에게는 뇌의 전두부의 혈류량이 다른 부분보다도
현저히 놓다. 반대쪽 (이 경우는 오른쪽) 손의 피부에 통증을 동반하지 않는
자극을 주어도 뇌의 혈류분포의 형태는 거의 변화지 않는다 (그림 7B 의 촉각자극). 즉 통각을 동반하는 자극 (그림 7B
의
통각자극) 에서는 혈류의 분포는 기본적으로 변하지 않지만 뇌의 총 혈류랑(그림 7B
의
각 뇌의 위에 % 로 나타냈다) 은 명백히 증가한다. 반대쪽 손을 자발적으로 열었다 닫았다
하면 (손의 운동) 총 혈류량은 증가한다. 이 경우에는 중심후회의 체성 감각영역과
그 부근의 두정피질의 혈류량이 동시에 증대된다. 이야기를 하거나 독서를
하면 혈류량의 증대부는 I 형의 분포로 되고 특히 독서 때에는 혈류의 중대부가 후두엽의
시각영역에까지 확대된다. 생각을 하거나 수를 세거나 하는 실험에서는 뇌의
총혈류량이 증가하고 중심구의 전 및 후부의 혈류량이 증대하게 된다.

<그림 7> 방사성 키세논가스
(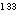 Xe) 의 동맥내 투여해 의한 뇌의 국소혈류량의 측정
Xe) 의 동맥내 투여해 의한 뇌의 국소혈류량의 측정
A: 방법
B: 안정 때와 활동 때에 있어서 언어중추 (좌) 측의 뇌혈류 분포, 안정 때의 뇌의 총혈류량을 100 % 로 해서 안정 때보다도 20% 이상 혈류가 증가한 부위 (적색원) 와 감소한 부위 (백색원) 을 나타낸다.
이상의 설명에서 밝혀진 것과 같이 감각, 운동, 사고 때 등 어떤 부분의 뇌가 활동한다 해도 뇌의 총혈류량이나 뇌의 국소혈류의 분포 또는 그 양쪽에 변화가 생긴다. 즉 뇌의 어느 부분의 신경활동이 상승하면 그 부분의 신경세포대사가 높아지고 산성대사 산물이 증가하면 뇌의 국소혈관 확장을 일으키고 그 결과 국소혈류량은 증가한다. 그반대 현상 또한 일어날 것이다. 에너지의 지속적인 공급없이는 고도의 신경활동이 요구되어도 신경원은 기능할 수 없다. 이것은 예를 들어 가장 고도로 추상적 또는 정신적 과정에 관련된 신경원일지라도 모든 신경원에 꼭 해당하는 것으로 생각된다. 이와 같은 추론은 의식소실, 혼수에 빠진 환자나 심한 치매 또는 정신분열환자에 대하여 행한 Ingvar 등의 관찰에 의해서도 지지된다. 그들은 환자들의 감각, 운동, 정신기능의 저하와 그 기능에 관여하는 뇌의 국소혈류량이나 뇌의 총혈류량의 저하 사이에 밝혀진 상관관계가 있다는 것을 보여주고 있다.
분명한 결론은 의식적 또는 무의식적인 뇌의 활동의 모든 것은 이들의 활동에 관여하는 신경회로가 작동하려는 상황에서 처음으로 가능하다. 하지만 이 결론은 마음과 신경활동이 서로 어떤 식으로 연결되어 있는가 하는 그 형태에 대하여는 설명할 수 없다. 현재 이와 같은 연결을 발견할 수 있는 결정적인 실험방법은 없다. 그러나 앞에서 설명한 실험에서 밝혀진 것과 같이 피험자의 언어나 행동을 측정한 정신활동에는 그에 상응하는 아주 특이적인 정신활동을 동반하고 그런 신경활동 없이 정신활동은 일어나지 않는다는 것은 명확하다. 예를 들면 임상적으로 죽은 사람이 숨을 되돌릴 때에 신경활동이 결여되어 있었을 시기를 생각해서 그 시기의 인식에 대해 설명할 수 있지만 이와 같은 보고는 전적으로 믿을 수 없다. 그 이유는 장애가 있는 뇌가 그 기능을 소실할 때, 또는 그 상태로부터 각성될 때에 많은 요소가 현재에 기초가 없는 지식 (환각) 을 일으키기 때문이다.
이와 같은 환자들에 의해 '사후의 세계'의 기록설명은 현저히 유사하다. 이와 같은 사실로부터 생각하면 모든 사람의 뇌는 구조가 공통이고 뇌의 에너지공급이 차단된 후 다시 에너지가 공급되어 그 기구가 회복될 때에 같은 모양으로 회복될 것이다. 예를 들면 텔레비전의 전원(에너지)을 켰다껐다 할 때에 어느 텔레비전 (기능) 에서도 같은 모양으로 빛이 커졌다꺼졌다 하는 것과 유사하다. 어떤 종류의 편두통이나 간질 때의 환시의 경우에도 아주 똑같이 일어난다. 안성 편두통 ophthalmc migration 의 환자는 종종 캄캄한 속에서 빛이 지그재그로 달리는 환각을 일으키는 것 (섬광성 암점) 을 알 수 있었다. 신비한 체험, 예를 들면 Bingen 의 성 Hildegard (1098 ~ 1179) 는 이와 같은 선명한 환각을 천국의 도시로 생각했을 것이다.
거의 모든 생물의 행동이나 기관의 상태는 주기적 변동을 나타낸다. 그 주기는 보통 24 시간의 낮 동안의 주기로 낮주기 diurnal rhythm 라 하고 이전 환경의 인자변화에 대해 생체가 몸에 받는 반응이라고 믿어왔다. 그러나 모든 환경인자를 일정하게 유지했을 때 조차 주기적 변동은 지속된다. 그러므로 주기성의 기본을 환경의 변화에서가 아니라 생물의 내인성 과정이라고 추정할 수 있다. 그 실체는 알려지지 않았지만 주기적 변동의 향도잡이 pacemaker 를 생물시계 biological clock 라고 한다.
일정한 환경조건하의 실험에서 계획된 생물 시계의 자유해방주기 free running period 는 보통 24 시간보다 다소 어긋나며 하루주기 (diurnal rhythm 을 낮주기로, circadian rhythm 을 하루주기로 구분했다) circadian rhythm 라고 한다. 자연조건하에서는 생체내의 하루주기는 외부동조신호 external entraining signal 에 의해 지구 자전의 24 시간 리듬과 같은 기간으로 하고 있다. 사람에 있어서는 하루주기의 동기화에 가장 큰 동조신호는 사회적 인자이고 주야의 명암교차와 같은 다른 인자는 그다지 중요하지 않다. 야근 노동자와 같이 어떤 사람의 생활 리듬이 일반 사회환경의 리듬과 다를 때에도 보통 일반 사회환경의 주기성은 그대로 남아 있다. 이 결과 특히 이른 아침 등의 어떤 일정한 시간에 동쪽이 둔해지는 것은 피하고 싶어하고 그래서 이와 같은 시간에 사고를 당하는 위험이 특히 높아진다.
사람의 낮주기는 실제로 많은 특정 장기 혹은 전신의 모든 기능에 영향을 미친다. 체온은 낮주기 변동을 나타내서 이른 아침에 최저값을 취하고 저녁에는 이보다 1 ~ 1.5 ℃ 높은 최고값을 취한다. 그러나 가장 현저한 낮주기 변동은 각성과 수면의 주기이다. 수면상태에 들어가기 전에 체온, 심박수, 호흡수의 저하 등의 수많은 조절이 이루어진다. 수면에 들어갈 때 이들의 기능변화가 수면의 원인이라고까지 생각되었다. 그러나 많은 생리학적 및 심리학적 여러 기능의 낮주기성은 수면을 끊어도 (수면차단 sleep deprivation) 그대로 지속된다. 그러므로 사람 (또는 다른 고도로 발달한 후생동물 metazoa) 은 다수의 생물시계 (발진기) 를 갖고 이들은 서로 같은 기간 같은 때에 외부동조신호에 의해서도 같은 기간에 이루어진다고 생각된다.
사람은 환경으로부터 격리되면 (예 : 지하의 움집이나 동굴에 격리) 하루주기 circadian rhythm 는 자유해방주기로 되고 그 주기는 24 시간 보다 약간 길어지는 경향이 있다. (그림 8A). 예를 들면 체온은 24시간 주기에서는 보통 각성기의 말기에 최고값 (그림 8A 의 적색 삼각표) 에 도달하지만 자유해방주기에서는 각성기의 초기에 이동한다. 이와 같은 사실로부터 정상 때 체온의 낮주기 diurnal rhythm 변동은 활동과 휴식의 리듬 결과로서 일어난 것이 아님을 알 수 있다. 오히려 이와 같은 관찰은 위에서 설명한 생각, 즉 체온의 주기와 활동의 주기가 각각 체내시계에 의해 지배되고 있다고 하는 생각과 일치하고 있다. 아래에 설명하는 것과 같이 체온과 생활의 주기는 전적으로 같은 기간이 아닌 것도 있다고 하는 관찰은 이 가정을 뒷받침하는 것이다.
<그림 8B> 에 나타난 실험에서는 15 일쯤에 윈인은 모르지만 돌연히 피검자의 활동주기가 33.4 시간으로 연장된다. 그러나 체온시계 temperature clock 는 활동시계 activity clock 보다도 명확히 유연성이 없고 33.4 시간이라고 하는 극단적으로 긴 활동주기란 같은 기간이 아니고 약 25 시간 주기를 유지한다.
이와 같은 각 체내 주기의 비동기화는 프랑스의 연구자도 보고하고 있다. 그는 외계로부터 완전히 격리된 동굴에 수주간에서 수개월을 보내면 피시험자의 수면-각성 주기는 종종 48 시간이 된다는 것을 보여주었다. 이와 같은 보통의 2 배의 14 시간 수면-34 시간 각성이라고 하는 리듬은 피험자에 있어서는 24 시간 주기로 보내고 있다고 생각하고 전적으로 정상인 것으로 느꼈다. 이와 같은 상태에 있어서 자율기능 (체온, 심박수, 호흡횟수 등) 은 긴 주기에서 전적으로 같은 시기로 하지 않고 독자적으로 25 시간 주기를 유지했다.

<그림 8> 사람의 하루주기
A: 문이 열려 있는 격리실 (즉 동조신호가 있는 상태) 과 문이 닫혀 있는 격리실 (동조신호가 없는상태) 에 있어서 피험자의 각성 (적색선) 과 수면 (청색선) 의 주기, 삼각표는 체온이 최고로 되는 시간을 나타낸다. 문이 열려 있는 방에서는 항상 정확히 24 시간 주기 (1 일의 평균편차가 ±0.7 과 ±0.5 시간) 를 나타냈다. 그러나 격리실에서는 주기는 26.1 ± 0.3 시간을 나타낸다.
B: 격리실에 있어서의
피험자의 활동주기
이 피험자가 체온주기 (최고값, 적색의
상향삼각형, 최소값, 회색하향 삼각형) 는 15 일째는 각성-수면
주기와 일치하지 않고 그후 25.1 시간주기를 유지한다. 15 일째에는
각성-수면주기 (활동주기) 는 돌연 33.4 시간으로 되지만 그 원인은 밝혀지지 않았다.
외부동조신호의 리듬에 1 회의 혼란이 생겼을 때 (예: 동쪽을 향해 비행하는데 하루의 길이가 단축되거나 서쪽으로 비행하는데 하루의 길이가 연장되는 것과 같은 경우) 하루주기 circadian rhythm 가 새로운 곳의 동조신호에 맞추어 올바른 위상으로 다시 같은 리듬으로 되기까지 여러 주기분의 시간이 걸린다. 재동기에 요하는 시간은 하루주기를 갖는 각종 기능에 따라 다른다. 이와 같은 리듬의 혼란은 jet lag 라고 하는 장거리 비행 후의 일시적인 능력저하의 원인으로 되고 있다.
하루주기의 생물학적 의의는 일반적으로 경시되어 왔다. 질병의 치료나 진단에 있어서도 여러 기능의 낮주기에 세심한 주의를 기울여야 한다. 하루주기성은 유전한다. 이것은 환경에 대한 계통발생적 적응현상으로 볼 수 있다. 실제로는 하루주기는 외계주기의 생체내 사본 internal copy 과 같은 것이다. 이와 같은 체내시계 internal schedule 의 도움에 의해 생체는 정확한 시간에 또는 미리 예상되는 환경변화에 적응할 수 있다. 예를 들면 체온과 많은 호르몬의 혈장 중 농도는 사람이 일어나기 약간 전에 상승하기 시작한다.
똑같이 동물에 있어서도 하루주기에는 많은 이점이 있다. 예를 들면 주기는 하루 중의 특정기간에 특정의 행동을 일으키는 것을 촉진한다. 극단적인 경우에는 체내시계는 시간의 측정에 사용된다. 이것은 태양을 나는 방향을 컴퍼스로 이용하는 동물에 있어서 필요한 능력이다. 이렇게 생각하면 각성-수면리듬은 원인이 아니라 오히려 내인성 하루주기의 외면적 표현의 하나에 지나지 않는다.
각성 중의 사람은 환경에 대하여 적극적으로 반응하고 자극에 대해서도 적절한 행동을 취한다. 수면 중에는 환경과의 접촉이 극도로 감소한다. 각성시 외부에 대한 주의가 여러 정도가 있는 것과 같이 수면 때에도 여러 가지 수면상 stage of sleep 이 구별된다. 단순 또는 고전적인 수면의 깊이 척도는 피험자를 깨어나게 하기 위해 필요한 자극의 강도이다. 깊은 수면에서는 자극의 역치가 높아진다.
오늘날에는 수면의 깊이 측정방법으로서 보통 뇌파를 이용한다. 뇌파로부터 4종류의 수면형태가 구별된다 (그림 9). 잠이 깊어질수록 개략적으로 뇌파는 느리게 한다 (동기화 경향).
곧 수면방추파 또는 K 복합파라고 하는 특수한 파형군이 출현한다 (그림 9A, C, D). 수면이 가장 깊은 상태 (그림 9E) 에서는 파형이 느리고 진폭이 큰 S 파가 출현한다.

<그림 9> 뇌파에 기초를 둔 사람의 수면상 분류 (A) 와 야간에 있어서 이들 상의 출현빈도 (B)
흔히 사용되는 2
개의 명명법 (A-E 및 W, 1~4) 을
나타낸다. 수면의 정도는 위로부터 아래로 향해 깊어진다. A 또는 W 상: 잠들기
직전의 졸린 상태, 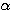 파가 우세하다. B(1) 상: 잠들 때
파가 우세하다. B(1) 상: 잠들 때 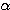 파는 소실되고 저진폭의
파는 소실되고 저진폭의 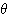 파가 출현한다. C(2) 상: 얕은 수면. 더욱 주파수가 감소해서
파가 출현한다. C(2) 상: 얕은 수면. 더욱 주파수가 감소해서 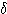 파, 12~15 ㎐ 의 수면방추파도 보인다. D(3) 상: 약간 깊은 수면,
파, 12~15 ㎐ 의 수면방추파도 보인다. D(3) 상: 약간 깊은 수면, 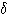 파 , K 복합파가 출현한다. E(4) 상: 깊은 잠, 뇌파는 큰 진폭의 느린
파 , K 복합파가 출현한다. E(4) 상: 깊은 잠, 뇌파는 큰 진폭의 느린 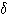 파가 우세하다. 뇌파에 관해서 REM 상은 B 상과 거의 일치한다. 어떤 상으로부터
다음 상으로 가는 것을 느리게 이행한다. 그림 A 의 뇌파는 기록지의 속도를 2 종류를
사용해서 기록했다 (시간축을 볼 것). B 는 3 일 밤을 연속해서 1 사람의 피험자의 뇌파를
측정하고 수면의 깊이 변동을 나타낸 것이다. REM상을 적색 막대선으로 나타냈다.
각 그림의 상부에 완전한 수면주기를 대괄호 (
파가 우세하다. 뇌파에 관해서 REM 상은 B 상과 거의 일치한다. 어떤 상으로부터
다음 상으로 가는 것을 느리게 이행한다. 그림 A 의 뇌파는 기록지의 속도를 2 종류를
사용해서 기록했다 (시간축을 볼 것). B 는 3 일 밤을 연속해서 1 사람의 피험자의 뇌파를
측정하고 수면의 깊이 변동을 나타낸 것이다. REM상을 적색 막대선으로 나타냈다.
각 그림의 상부에 완전한 수면주기를 대괄호 ( ![]() ) 로나타냈다. 그래프의 아래 종선은 몸의 움직임의 횟수가 강도
) 로나타냈다. 그래프의 아래 종선은 몸의 움직임의 횟수가 강도
사람은 야간의 수면 중에 여러 가지 수면형태를 평균 3 ~ 5 회 반복된다 (그림 9B, 10). 일반적으로 아침이 가까워짐에 따라 각각의 주기 중에 도달하는 수면의 최대 깊이는 감소하고 마침내 수면의 E형태는 일어나게 된다. 낮주기성을 나타내는 많은 자율기능 중에는 체온 등과 같이 수면깊이의 주기적 변동의 영향을 받지 않는 것이 있다. 한편 심박수나 호흡등에서는 낮주기에 있어서 수면의 깊이 변동에 일치하는 일시적인 변동이 겹친다 (그림 10). 이와 같은 일시적인 성분은 특히 수면의 B 형태에 동반해 출현한다. 수면 중에 B 형태에 일치해서 반복 출현하는 반응 (음경발기 등) 도 보인다 (그림 10).

<그림 10> 야간에 있어서 수면상과 그에 따른 자율기능의
여러 변화
평균적인 모식도로 나타냈다. a. 적색 곡선은 뇌파에 기초를 둔 수면의 깊이 변화를 나타낸다. REM상은 적색 막대선으로 나타냈다. b. 의 전기안구 그림에서 급속안구운동 (세로선으로 나타냈다) 은 REM 상 때에만 일어나는 것을 알 수 있다. 그 외에도 졸리운 때에 REM 약간의 안구운동 (b 그림 왼쪽 끝) 이 인정될 뿐이다. 목근육의 근전도를 C 에, 1 분간 심박수를 d에 나타냈다. REM 상 때 심박수는 일과성으로 증가한다. e 에 나타낸 호흡수도 REM 상에서 증가한다. 남성의 음경발기는 REM 층 때에 규칙적으로 일어난다.
운동계의 활동성도 수면 중의 B형태와 일치하여 반복해서 변한다. B 형태에서는 근육긴장은 깊은 수면 때와 똑같이 거의 소실된다 (그림 10, 근전도: EMG). 그렇지만 안구는 B형태에서 급속히 움직인다 (그림 10, 전기안구도 : EOG). 이와 같은 빠른 안구운동은 B 형태의 특징적인 것으로 B 형태를 REM rapid eyemovement 형태라고 한다. 각성시키는 데 필요한 외부자극강도 역치는 REM 수면 때에는 깊은 수면 때와 같은 높이이다. 그러나 REM형태의 뇌파는 수면에 들어간 직후의 것과 유사하다. REM 수면은 이와 같이 모순된 징후를 나타내기 때문에 모순수면 paradoxical sleep 또는 탈동기수면 desynchronized sleep이라고도 한다. 여기에 대하여 다른 수면 형태를 NREM 수면 또는 non-REM 수면이라고 한다. 보통 REM 형태는 수면 중에 약 1.5 시간마다 출현한다 (그림 9). 평균 약 20 분간 지속하지만 그 지속시간은 밤이 깊어질수록 연장된다.
나이와 더불어 하루에 상대적 수면시간은 감소하고
수면시간 중에서 REM 수면이 차지하는 비율도 감소한다. 이와 같은 경향을 <그림
11> 에 나타냈다. <그림 11> 에는 나타나 있지 않지만 수면의 각 형태의 순서라든가
지속시간은 어린이나 유아 그리고 성인에서는 확실히 다르다. 젖먹이 유아에서
REM수면이 높은 비율로 출현하는 것은 REM형태의 신경 활동항진이 어린 뇌의 성숙에
중요하다는 것을 나타내는 있다 (REM 형태의 EEG는 주의집중 때의 EEG 와 유사해서 탈동기되어
있다. <그림 9 의 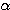 파차단>. 젖먹이 유아에서는 외부로부터의 자극효과가 비교적 적다. 또한 REM
수면
중에 특히 꿈 (꿈은 대뇌의식활동의 특수한 형태이다) 을 잘 꾸지만 이것도 REM 수면의
어린 뇌의 성숙에 대한 중요성을 나타내고 있다.
파차단>. 젖먹이 유아에서는 외부로부터의 자극효과가 비교적 적다. 또한 REM
수면
중에 특히 꿈 (꿈은 대뇌의식활동의 특수한 형태이다) 을 잘 꾸지만 이것도 REM 수면의
어린 뇌의 성숙에 대한 중요성을 나타내고 있다.

<그림 11> 수면 상태
A: 사람의 생애의 있어서 각성과 수면에 차지하는 평균시간과 REM 수면 및 비REM 수면의 비율, 수면시간의 감소는 각각으로 가장 현저한 변화는 어린이가 성장할 때, REM 수면 시간이 확실히 감소하는 것을 알 수 있다.
B: 여러 단계의 수면 동안 뇌의 전기적 활동을 뇌파로 기록했다.
C: 하룻밤 수면 동안 3~5 주기를 가지며 각 주기는 여러 가지 연속된 수면단계가 포함된다.
피험자의 노파가 REM 형태를 나타낼 때에 외부의
자극으로 눈을 뜨게 하면 피험자는 막꿈을 꾸었다고 하는 일이 많다 (60 ~ 90 %).
non-REM
수면형태에서는
눈을 뜨게 해도 꿈을 꾸었다는 일이 적고 동시에 연구자에 의한 혼란도 있다 (1 ~
74 %),
지금까지의 연구결과에 의하면 REM 수면을 단순히 꿈을 꾸기 위한 수면으로 간주하는
것은 옳지 않지만 REM 수면의 지속중에 꿈을 많이 꾸게 되는 것은 사실이다. 이와
같은 관점에서 REM 수면에 따른 여러 가지 기관의 기능변화 (그림 10) 를 꿈에 관련한
생리적 현상이라고 할 수 있다.
REM 수면과 꿈의 관계에 대한 추론을 하는 것은 피험자에 의한 꿈의 주고나적 경험의 보고에 의존한다는 점에서 방법론적으로 문제가 있다. 실험자의 질문방법론적으로 문제가 있다. 실험자의 질문방법에 따라 피험자의 대답도 영향을 받는다. 다른 한편 꿈과 REM 수면 사이에는 엄밀한 관계가 없다고 생각하는 관찰도 있다. 예를 들면 신생아도 <그림 11> 에 나타난 것과 같이 급속히 눈을 움직이는 것을 동반하는 수면의 형태를 갖지만 대뇌 피질의 발달이 이른 단계에서 시각적인 꿈의 경험이 있다고 생각하기 어렵다.
이같은 경우가 다른 포유동물의 신생아에도
해당된다. 더욱이 선천적인 맹인에서도 빠른 안구운동이 관찰되었지만 당연히 이
사람들은 시각의 경험이 없다. 더욱이 만성 제뇌고양이에서조차도 REM 형태의 특징적인
말초의 효과가 반응을 갖춘 수면형태를 취한다. 이와 같은 사실로 보아 REM 수면의
모든 징후는 꿈을 꾸는 결과로 생기는 2 차적인 현상이 아니고 오히려 성숙한 뇌에서는
꿈은 REM 형태의 출현 때에 일치해서 일어난다고 하는 쪽이 적절할 것이다. 사람에서
REM 수면상태가 개시될 때에 반드시 외부로부터 눈을 뜨게 해서 소위 REM 수면차단
REM-sleep deprivation 상태로 하면 그후의 중단되지 않은 수면에서는 REM
수면형태의 비율이 거의 50 ~ 80 %
상승한다. REM 수면차단을 16일 이상 반복하면 전체의 수먼이 약간 단축되어 버리는
것의 영향에도 의하지만 피험자는 짜증을 내게 되고 더구나 눈을 떴을 때나 한번
잠을 깨는 것과 같은 행동을 취할 때조차 짧은 REM 수면형태가 일어난다. 피험자는
종종환각상태에서 고통스러워한다.
수면차단으로 인한 이상의 증상 중 어느 부분이 꿈을꾸게
하는 것에 기인하는지를 명확하지 않다. 그러나 수면차단을 행한 피험자에 있어서
REM 수면차단, non-REM 수면차단, 혹은 양쪽 수면의 차단 어떤 것에도 장기간의 중증의
정신적 또는 신체적장애는 인정되지 않는 것을 부연해 둔다.
사람이나 동물의 일상생활의 수면의 강한 욕구가 주기적으로 일어나는 것으로부터 다음의 가장 그럴 듯한 해석이 시사된다. 결국 피로와 수면은 혈액 중의 대사물질의 주기적인 생산, 축적에 주로 기인하고 수면 중에 이들 대사산물은 제거, 분해 또는 다른물질로 치환된다고 하는 것이다. 물론 이와 같은 각성과 수면의 화학설을 단순히 지나쳐 버리는 것은 옳지 않다. 이와 같은 물질 존재의 시증 자체, 아직 얻지 못한 것은 물론 화학설에 대한 결정적 반증이 나타나고 있다. 순환계를 공유하고 있지만 신경계는 각각 독립되어 잇는 Siamese 쌍생아에 있어서 수면과 각성의 주기는 서로 영향을 주지 않고 두 사람 각각에 일어나고 있다. 그외에도 대뇌피질만을 분리한 동물이라든가 양쪽의 대뇌반구를 정중시상면으로 절단 분리한 동물에서 수면이나 각성은 절단된 뇌의 각 부분에 있어서 독립적으로 일어난다.
각성과 수면주기를 이해하기 위해 각성과 수면중 뇌의 신경활동이 다른 것을 추구하는 연구가 행해지고 있고 수면상태와 각성상태 사이의 이행을 가져오는 신경과정을 알려는 시도가 이루어지고 있다. 대단히 중요한 것이지만 수면은 각성 때문에 보이는 뇌의 신경활동 형태의 단순한 결핍 또는 뇌의 일종의 휴식은 아니다. 신경생리학적 자료로부터 각 수면상태에 있어서 신경활동은 각성 때와 똑같이 복잡하다는 것이 증명 되고 있다. 예를 들면 여러 가지 수면형태 중 뇌파의 변동 정도 (그림 5, 9) 는 각성 중의 것과 똑같은 정도이다. 꿈의 존재도 대뇌가 수면 중에 쉬지 않고 있다는 것을 싫증하고 잇다. 수면이란 대뇌의 작용양식이 변화하는 것을 의미하고 있고 통합적 신경활동의 결핍을 의미하는 것은 아니다. 신경활동의 결핍은 깊은 마취나 혼수에서 일어난다.
뇌간은 각성과 수면의 억제중추 요소이다. 뇌간망상체에
고빈도의 전기자극을 주면 수면 중의 동물은 각성되고 뇌파의 탈동기가 일어난다.
역으로 이 부분의 파괴 또는 이곳에서 피질로 올라가는 경로의 절단 (이 경로는 감각
등의 특수경로와 구별되기 때문에 비특수투사 nonspecific
projection 라고 한다) 에 의해 동물은 수면에 빠진다. 이와 같은 발견에 의해 '수면과 각성이
망상체설' 이 생겼다. 이 이론에 의하면 망상체에는 상행성의 활성화 흥분충동에 의해
각성상태의 흥분수준을 만드는 기능이다. 이와 같은 계통을 상행성 망상체활성계통
ascending reticular activating
system : 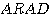 이라고 한다. 상행성 망상체활성계통의 활동변동으로 수면으로부터 각성으로 또는
그 반대의 이행이 일어난다고 생각된다. 각성상태에 있어서 미묘한 행동의 변화,
예를 들면 주의깊은 정도의 변화 등은 ARAS 의 활성의 미세한 변동에 의한다고 생각된다.
이와 같은 모양의 망상체를 각성의 중추로 하는 단순한 설은 최근 수많은 발견과
일치하지 않고 적어도 이 설의 원형대로는 받아들여지지 않는다. 예를 들면 망상체가
파괴된 뇌에서도 수면과 각성의 리듬을 나타내고 망상체는 그 리듬에 불가결한 것은
아니다.
이라고 한다. 상행성 망상체활성계통의 활동변동으로 수면으로부터 각성으로 또는
그 반대의 이행이 일어난다고 생각된다. 각성상태에 있어서 미묘한 행동의 변화,
예를 들면 주의깊은 정도의 변화 등은 ARAS 의 활성의 미세한 변동에 의한다고 생각된다.
이와 같은 모양의 망상체를 각성의 중추로 하는 단순한 설은 최근 수많은 발견과
일치하지 않고 적어도 이 설의 원형대로는 받아들여지지 않는다. 예를 들면 망상체가
파괴된 뇌에서도 수면과 각성의 리듬을 나타내고 망상체는 그 리듬에 불가결한 것은
아니다.
수면과 각성현상에 있어서 뇌간의 중추적 역할은 신경약리학적 연구에서도 강조되고 있다.수면과 각성의 주기에서 특정의 뇌간핵으로부터 monoamine 전달물질인 serotonin (5-HT) 과 noradrenaline 의 방출이 정기적으로 생긴다. 수면과 각성의 생화학설은 차례로 확립되어가지만 다음의 동물실험에서의 관찰사항은 특히 중요하다.
① 뇌간내의 봉선핵 raphe nuclei 의 신경원은 다량의 serotonin 을 함유하고 있다. 이 serotonin이 고갈되면 (예: serotonin 합성과정의 저해) REM 및 non-REM 수면 모두에 영향을 받고 중증의 불면증 insomnia 으로 된다. 그 불면증은 serotonin 의 전구물질인 5-hydroxytry-ptophane 의 투여에 의해 완화된다(serotonin 은 혈액뇌장벽을 통과할 수 없다).
② 뇌간내에 청반 locus ceruleus의 신경원에 다량의 noradrenaline이 함유되어 있다. 이 청반을 양측성을로 파괴하면 REM수면으 소실되지만 non-REM수면에는 영향이 보이지 않는다
③ Serotenin 과 noradrenaline 을 reserpine 의 투여해 의해 고갈시키면 non-REM 수면, REM 수면 모두 소실된다. 그 결과 동물은 중증의 불면증에 빠진다. (상기 ① 의 성적으로부터 기대되는 모양으로), 이어 5-hydoxytrytophane 의 투약으로 non-REM 수면은 회복되지만 REM 수면은 소실된 그대로다. 이것은 상기 ② 에서의 관찰과 일치한다.
이와 같은 소견은 2 종류의 물질이 수면에 밀접히 관계한다는 것을 시사한다. serotonin 은 non-REM 수면에, noradrenaline 은 REM 수면에 관계하고 보통 REM 수면은 non-REM 수면에 선행된 때에만 출현 가능하다.
그러나 사람의 연구에서는 sertonin 과 noradrenaline 의 반대의 관여가 시사되고 있다. 즉 REM 수면은 오히려 serotonin 이 증가할수록 그리고 noradrenaline 이 감소할수록 많아진다고 한다. 이와 같이 모순된 원인은 해명되지 않고 있다.
잠에서 깨어났을 때의 의식이 점차 회복될수록 일어나는 현저한 변화를 우리들은 달리 체험하지 못한다. 의식의 미묘한 변화는 내성적으로만 평가될 수 있는 것이지만 의식은 틀림없는 인간존재의 기본이 된다. 의식을 생리학적 및 심리학적으로 정의하려고 하는 수많은 시도가 있었다. 그러나 현재 의식의 정의는 일정하지 않다. 생리학에서는 자연과학의 입장에서 의식의 존재를 밝히려고 하는 것이 가능하다고 생각되는 경계조건 boundary condition 을 설정해서 의식을 객관적으로 생각한다. 사람과 동물의 의식존재의 지료로 되는 행동을 얻는 특징으로 다음과 같은 것이 있다.
① 주의깊이 및 합목적적으로 주의를 향하는 능력을 잦는다
② 추상적인 개념을 창조하고 동시에 사용할 수 있고, 그것을 언어 또는 다른 기호로 표현하는 능력을 갖는다.
③ 행위의 의미를 사전에 평가할 수 있고, 예상 계획하는 능력을 갖는다.
④ 자기와 타인을 인식하는 능력을 갖는다.
⑤ 미적 및 윤리적 평가능력을 갖는다.
이들 특징은 물론 의식의 기준으로 된 중요성에서 각각 다르고 그 중에는 언어와 같이 주로 사람에게만 볼 수 있는 것도 있다. 만약 이것이 의식의 존재를 나타내는 행동상의 특징으로 받아들여지게 되면 동물에도 의식이 있다고 하는 흥미있는 문제를 함유하고 있는 것으로 생각된다. 모든 동물이란 한계가 없지만 고도로 분화된 신경계통을 갖는 고등동물 (예: 조류나 포유류)은 위에서 설명한 행동상의 특징 중의 몇 개를 갖는다. 단순한 신경계통을 갖는 동물에 있어서는 드물 게 존재하는 경우도 있다. 이와 같은 행동상의 특징으로부터 정의한 특징은 복잡한 신경구조와 밀접한 관계가 있다.
위와 같은 것에 의해 의식을 갖는 동물과 갖지 않는 동물 사이의 명확한 경계는 존재하지 않는다는 것이 확실하다고 생각된다. 의식은 신경계통의 계통발생학적인 발달과정과 거의 비례해서 발달하는 것 같이 생각된다. 다시 말하면 동물계에는 여러 가지 발달단계 또는 여러 형태의 의식이 있고 그래서 그 중 사람의 의식이 의심할 것 없이 가장 분화된 형태일 것이다.
의식을 갖는 데는 적절히 분화된 신경계통을 필요로 한다고 하는 견해를 기초로 다음과 같이 유추할 수 있다. 결국 진화 (계통발생)의 과정에 있어서 단순한 형태의 신경활동 (예: 반사) 에서는 생체의 조절이나 관리가 충분히 이루어지지 않았을 것이고, 그랬을 때에 의식이라고 하는 것이 어떤 형태로 출현했던 것이라 여겨진다. 만약 그렇다면 의식의 발달은 필연적인 진화과정이고 고등동물이 환경에 잘 적응하기 위한 불가결한 방법이었을 것이다.
사람 의식의 기능적 필요조건, 결국 뒷받침되는 신경활동에 관한 지식은 현재 불충분하다. 의식은 각성 때의 탈동기화 뇌파에서 나타난 것과 같이 중간 정도의 신경활동에 어울린다. 의식은 신경활동이 전기 쇼크나 간질발작과 같이 항진되어 있을 때나 마취나 혼수일 경우와 같이 항진되어 있을 때나 마취나 혼수일 경우와 같이 저하되어 있을 때에는 유지되지 않는다. 의식은 또한 피질과 피질하와의 상호작용의 기초에서만 존재가능하고 그 어느 부분도 단독으로는 의식을 유지할 수 없다.
최근 뇌량 (양쪽의 대뇌반구를 연결하는 교련섬유가 통하는 부위)의 절단수술을 받은 양단뇌 split - brain 환자의 소견에서 의식이 신경구조에 관한 중요한 보고가 계속되었다. 이들 환자에서는 <그림 12> 에 나타난 것과 같이 뇌내에서 교차하는 다수의 상행성 및 하행성 경로에 의해 대뇌의 좌반구는 우측반신의 운동 및 체성 감각을, 우반구는 좌측반신의 여러 가지를 지배한다. 시신경도 시교차를 개재해서 시야의 우반분이 대뇌의 좌반구에 투사되고 좌반분이 우반구에 투사된다. 다른 한편으로 중추성 청각로는 일부 교차하고 일부는 교차하지 않기 때문에 각 반구는 양쪽 귀에서 오는 입력을 받는다.
양단뇌환자의 수술 후 상태는 상기의 각 기능을 미묘하게 반영한다. 이 환자에서는 지능을 포함한 일상생활을 영위하는데 현저한 변화는 보이지 않는다. 환자가 오른손잡이의 경우는 좌반신의 자발적인 움직임이 약간 감소하고 좌반신으로 오는 자극에 대한 반응에 약간의 저항을 느끼는 정도이다.

<그림
12> 양단뇌환자에서 볼 수 있는 체성 감각, 운동, 시각, 청각의 섬유 연락
좌반구는 우반신만 그리고 체성 감각성 (구심성) 및 운동성 (원심성) 연락을 한다. 좌우 눈의 시야의 오른쪽 (왼쪽) 반분은 좌 (우) 반구의 시각피질로 투시된다. 대조적으로 좌우의 귀는 양단뇌환자에 있어서도 양쪽 뇌반구의 청각피질과 연락한다.
그런데 <그림 13> 에 나타난 것과 같은 실험장치를 쓰면 분리된 양측의 대뇌반구가 상당히 다른 역할과 능력을 갖고 있다는 것이 밝혀졌다. 이 장치에서는 시야의 우반분과 좌반분에 별도로 시각신호 (섬광, 문자) 를 보낼 수 있고 한쪽의 대뇌반구만이 신호를 받는 것이 가능하다. 더욱이 실험대 위에 놓인 물체를 환자에게 보이지 않게 좌우 어느 손으로 만져서 판단하거나 글씨를 쓸 수 있도록 한다. 이 경우에도 <그림 12> 와 일치하고 오른손 (왼손) 의 운동신경 및 감각신경은 좌반구 (우반구) 와만 연락하고 있다. 위에 설명한 실험의 중요한 소견을 다음에 요약한다.
.gif)
<그림
13> 양단뇌환자 실험(I)
Sperry 등의 공동연구자가 양단뇌환자의 검사에 쓰인 실험장치를 나타냈다. 환자는 그림이나 문자가 투영되는 반투명 스크린 앞에 앉게 한다. 그림이나 문자는 왼쪽의 시야, 오른쪽의 시야 또는 양쪽에 나타나는 것처럼 된다. 환자는 스크린 중앙의 점을 응시하도록 한다. 다른 시야의 자극을 막기 위해 시각자극이 단기간 (0.1초) 주어지고 이때 즉시 방향이 변하지 않는 모양으로 된다. 환자는 스크린 아래로 손을 뻗쳐 시각을 억제하지 않고 손 일을 할 수 있다. 이것은 필름에 기록된다. 청각 신호는 이어폰을 통해 주어지고 실험환자에도 들린다.
양단뇌환자는 어떤 특정한 물체를 시야의 우반분에만 보여주면 그 명칭 (예: 열쇠, 연필 등) 을 대거나 오른손에서 그 물체를 많은 물체 중에서 선택해 낸다. 어떤 단어를 시야의 우반분에 투사시키면 환자는 그것을 읽고 쓰고 오른손으로 해당하는 물체를 선택할 수 있다. 그 특정의 물체를 환자의 오른손에 쥐어 주었을 때에도 환자는 그 명칭을 말하고 기술할 수 있다. 결국 환자는 이 조건하에서는 정상 피험자와 똑같이 행동한다.
그런데 시야의 좌반분에만 특정한 물체를 보여주면 양단뇌환자는 그 명칭을 대지 못한다. 환자에게 왼손으로 다른 물체 중에서 동일물체를 선택해 내라고 명령하면 환자는 올바르게 선택해 내지만 선택 후에도 그 명칭을 대지 못한다. 왼손에 물체를 쥐어주었을 때에도 환자는 그 명칭을 대지 못했다. 단어를 시야의 좌반분에 보여주면 환자는 그것을 소리내서 읽지 못하지만 단어가 일용품의 명칭이면 왼손으로 올바로 물체를 선택한다 (그림 14). 이와 같은 경우에도 올바로 물체를 선택한 후에까지도 그 명칭을 대지 못한다. 이와 같은 실험조건하에서 환자는 더욱 고도의 작업도 실행할 수 있도록 명령을 해도 말로 설명하거나 써서 설명할 수 없다. 결국 환자는 우반구를 작용시켜서 실행한 것은 전적으로 제멋대로 행동한다.
위에 설명한 성적으로부터 다음과 같은 결론을 얻을 수 있다. 좌우로 분리된 대뇌의 좌반구 능력은 환자의 자각이나 일상의 행동이라든가 위에서 설명한 실험에 대한 반응으로부터 손상 없는 뇌의 능력과 구별되지 않는다. 이것은 특히 환자가 내성적으로 체험하는 의식 및 이것에 대하여 환자자신이 설명하는 것으로도 증명된다. 따라서 정상 뇌에 있어서도 좌반구를 그것과 연락이 있는 피질하의 부분과 함께 사람에 특유한 의식과 그것에 따르는 언어능력의 기초를 이루는 신경구조로 보아야 한다.
.gif)
<그림 14> 양단뇌환자 실험 (Ⅱ)
Sperry 등의 공동연구자에 의해 실험중에 보여준 양단뇌환자의 행동, 환자는 왼쪽의 언어반구를 써서 오른쪽 시야의 '반지' 라고 하는 단어를 읽었다고 보고했다. 환자는 왼쪽 시야의 '열쇠' 라는 단어를 본 것을 부정하고 또한 왼쪽손에 놓인 물체의 명칭도설명할 수 없다. 그러나 물체의 명칭은 모르지만 왼쪽 손으로 옳게 물체를 선택할 수 있다. 역시 선택된 물체의 명칭을 들으면 언어 반구의 작용에 의해 '반지' 라고 대답할 것이다.
좌반구에서 분리된 대뇌의 우반구는 언어에 의해 자신을 표현할 수 없다. 우반구에서 행하는 감각처리, 통합작용, 운동과정 등은 확실하게 환자의 의식에 도달하지 못한다. 우반구는 좌반구로부터 분리되면 독립적으로 작용하고 환자는 우반구의 작용에 대해 좌반구의 감각계통을 통해 간접적으로만 알 수 있다. 동시에 우반구는 현저히 능력을 갖추고 있고 기억, 시각 및 촉각에 의한 형태의 인식, 추상화, 어느 정도의 언어 이해 등이 가능하다 (음성으로 내린 명령을 실행해서 얻는 간단한 언어도 이해할 것이다).
예를 들면 음악적 이해나 공간적 개념을 처리하는 능력 등의 몇몇 점에서 우반구의 능력은 좌반구의 그것을 능가하고 있다. 대체로 사람의 우반구의 능력은 유인원 (예: 침팬지) 을 포함한 다른 동물의 분리되지 않은 양반구의 능력을 상회하고 있다. 앞에서 설명한 고등동물의 의식문제로 되돌아오면 이미 설명한 행동상의 특징으로부터 이해할 수 있듯이 분리된 우반구의 의식은 고도로 발달되어 있다. 그러나 우반구는 하등동물과 똑같이 우반구의 의식상태를 언어로 표현 전달할 수 없다.
우반구의 의식 성질에 대해 아래의 사실로부터 더욱 짐작할 수 있다. 양단뇌환자는 한결같이 수술 후 꿈을 꾸지 않는다고 호소한다. 사실 수술 후 꿈을 꾸지 않는다고 호소한다. 사실 수술 후 환자의 수면 때의 좌반구는 뇌파는 이미 REM 형태를 나타내지 않는다. 그런데 우반구의 수면 때 뇌파는 여전히 명확한 REM형태를 나타낸다. 즉 우반구에서만 꿈을 꾼다고 하는 경험은 있을 수 없지만 우반구는 수면 중, 꿈을 꿀 것이다. 이러한 사실로 볼 때 정상상태에서 꿈은 뇌의 우반구에서 시작해 뇌량을 통해 좌반구로 확산된다고 생각한다.
상기 설명의 양단뇌환자의 연구에 의하면 언어능력을
떠맡고 있는 뇌의 영역은 원칙적으로 좌반구에만 존재한다. 이와 같은 것은 뇌손상환자의
임상적 관찰로부터도 옛날부터 추론되고 있다. 이와 같은 이유로 인해 좌반구를 우위반구
dominant hemispheredominant
hemisphere 라고 한다. 그러나 양단뇌환자의 연구로부터 우반구의 능력이 특수한 면에서 좌반구의
능력보다도 우수하다고 하는 증거가 축적되고 있는 중이다. 따라서 정확히 고찰해
보면 양쪽의 대뇌반구는 서로 보충하는 모양으로 특수화되고 보통 좌반구가 언어우위
language-dominant
로 되어 있다고 표현한다.
대뇌좌반구의 피질 중에서도 구별되고 언어에 관여하는 중요한 영역이 구별되며 그 영역을 언어영역 speech area,이라고 한다. <그림 15A> 에 전두엽 언어영역 frontal speech area, 측두엽 언어영역 temporal speech area, 3 차 언어 영역 tertiary speech area, 을 나타낸다. 위의 3 개 언어영역에 공통된 성질은 이 부위를 전기자극 (치료상 필요한 수술대에 국소마취된 환자의 협력하에 이루어진다) 하면 환자는 자극 중 실어를 일으키고 단어나 글을 이야기하지 못하게 되었다. 이 부분의 자극에 의해 말을 하는 것은 없다.

<그림15> 언어우위 (좌) 반구의 언어영역 (A, 적색) 과 비언어우위 (우) 반구의 상당한 영역 (B)
Penfield등의 공동연구자에 의해 성인환자의 노출된 피질을 전기자극함으로써 확립되었다. 발성에 관여하는 근육의 제어 영역은 대뇌 양쪽의 중심전회에 국재한다. 다른 신체의 피질운동제어란 현저히 다르고 회화에 쓰이는 좌우의 근육조직은 둘다 양측성으로 제어된다.
Broca 가 백여년 전에 전두엽의 언어 영역 손상에 의해 언어능력이 장애되는 것을 지적한 이래 전두엽 언어영역을 Broca 영역이라고도 한다. 이 부분의 손상을 받은 환자는 언어를 이해하는 능력을 유지하지만 자발적으로 거의 말을 할 수 없고 명령된 짧은 글을 말하는 것조차 곤란하다. 이와 같은 상태를 운동성 실어증 motor aphasia 이라 하고 그때 발성에 필요한 골격근군은 건전하고 음식물 섭취도 정상으로 이루어진다.
Wernick 는 앞에서 설명한 Broca의 발견과 같은 시대에 측두엽의 언어영역이 파괴되면 언어이해, 즉 수용면이 매우 깊이 장애되는 것을 발견했다. 측두엽 언어영역을 Wernicke 영역이라고 한다. 이 영역이 장애된 상태를 감각성 실어증 sensory aphasia 이라고 한다. 이영역이 손상되면 전두엽 언어영역이나 3 차 언어영역 (Penfield 등이 앞에 설명한 수술 중의 자극실험에서 짧은 기간의 실어가 일어나는 것으로부터 발견했다)의 손상 때에 비해 매우 위독한 것으로 장기간의 (때로는 영구히) 장애가 일어난다.
Broca 와 Wernick 가 자기가 발견한 영역에 있다고 생각한 기능은 각각 과도하게 단순화된 것이고 1 차적으로 근사한 것이지만 옳지 않다. 결국 표현성 (운동성) 및 수용성 (감각성) 언어기능과 거기에 부수된 읽고 쓰기나 계산의 여러 기능은 결코 일정한 형태로 장애되는 것은 아니고 여러 가지 다른 것들이 짜 맞추어져서 장애된다. 따라서 최근의 임상적 및 심리학적 입장으로는 여러 가지 형의 실어증을 뇌의 특정영역에 결부시키는 것이 불가능하다고 생각하고 있다. 본장에서는 각종의 실어증이나 각각의 특징과 분류에 대한 최근의 상세한 성과에 대하여는 설명하지 않는다.
실제로 오른손잡이 사람의 경우, 언어중추는 좌반구는 있다. 의외의 것으로 왼손잡이 사람도 보통도 좌반구에 있지만 드물 게 우측이나 양측에 있는 경우도 있다. 반구의 어느 쪽에 언어중추가 존재하는가를 조사하는 간단한 실험이 있다 초단시간마취약 (barbiturate amytal) 을 측경 부위의 한쪽 총경동맥으로 주입하면 그쪽의 반구만이 곧 마취된다. 반대쪽은 각성된 채로 있다. 이때 환자가 시험자와 이야기할 수 있으면 각성쪽의 반구에 언어중추가 있고, 할 수 없으면 반대쪽에 있지만 드물 게 양쪽에 언어중추를 갖는 사람이 발견되는 일이 있다. 이 실험은 주로 곤란한 뇌수술 때에 수술에 의해 환자의 이야기 능력이 장애되지 않는 것을 확인하기 위해 예비 실험으로 행해진다.
이야기하는 것을 깨달은 어린이 뇌의 좌반구에 있는 언어영역이 사고로 장애되면 전적으로 이야기하지 못하게 되지만 1년쯤 경과하면 어린이는 다시 이야기를 시작한다. 이 경우 이야기기능은 대뇌의 우반구의 대응하는 부분이다 (그림 15). 좌반구로부터 우반구로 오는 이와 같은 언어우위성의 전이는 뇌의 경이적 능력의 하나이고 뇌의 기질적 장애 후에 적응하는 이와 같은 능력을 가역성이라고 한다.
좌반구로부터 우반구로 오는 언어우위성의 전이는 늦어도 약 10세까지의 한정된 시기로, 그 이후는 일어나지 않는다. 이 이상의 나이가 되면 이와 같은 전이는 아마 아래 두 개의 이유로부터 일어나지 못하게 된다. 하나는 말하는데 필요한 기본적 신경원회로 (외국어를 공부할 때에도 활용된다) 의 형성이 이 시기를 지나면 불가능하게 되고, 또 다른 하나는 언어우위가 차지해야 할 반구의 대응하는 부분이 이 시기 이후에는 주로 자세의 공간적 제어를 포함한 공간적인식 (목표의식) 과 같은 다른 기능에 의해 차지 되기 때문이다.
위에서 설명한 뇌의 가역성이 활용될 때에는
유리한 면뿐만 아니라 한 편으로는 불리한 면을 동반한다. 유아기에 좌반구에 손상을
받아 우반구가 언어뇌로 되어 있는 환자에서는 언어능력이 평균보다 열악할 뿐만
아니라 일반적으로 지능도 낮다.
좌반구 언어영역에 대응하는 부분인 우반구의 두정엽 및 측두엽 (그림 15 의 A 와B 를 비교해 보라) 의 손상은 공간실인 spacial agnosia 이라고 하는 공간적 목표의식 장애를 일으킨다. 증상은 여러 가지이지만 예를 들면 일상의 환경에서도 자기가 사는 장소의 위치나 지리를 잃고 입방체 등의 간단한 형태의 입체그림이 그려지지 않는다.
생체를 환경에 적응시키는 신경계통의 일반적 특성으로서 정보의 수용, 유지 및 재생의 기능이 있다. 학습, 기억 또는 정보의 재생능력이 없으면 계획성을 가지고 성공하거나 의식적으로 실패를 피하는 것이 되지 않는다. 최근 이런 문제가 신경생리학자나 심리학을 시작하고자 하는 다른 분야의 연구자가 관심을 모으게 되었지만 충분히 만족할 만한 설명은 없다. 또한 학습과 기억의 연구에 비해 재생기능의 연구는 거의 이루어지지 않는다.
사람의 기억에 대해서 확립된 실험사실은 적지만 확실한 것은 의식으로 취급되는 사항의 일부분만이 장기적으로 기억될 수 있다는 것이다. 의식에 관여하는 정보는 감각정보 전체의 일부에 지나지 않지만 의식정보의 1 % 만이 선택되어 장기기억으로 남는다. 더구나 사람은 기억한 정보의 대부분을 망각한다. 이와 같은 선택과 망각의 2 개의 기구에 의해 사람은 학습과 기억의 결여와 같은 정도로 유해한 정보가 범람하는 상황으로부터 피할 수 있다.
다음으로 중요한 사실은 짧은 사항 (예: 의미가 없는 음절의 열거 등) 의 기억방법이 긴 사항의 기억보다 용이한 것이다. 이 사실은 당연한 것으로 생각되지만 기억이 자기 (磁氣) 테이프같이 용량을 재거나 정지명렬이 나올 때까지 정보를 계속 기록하는 것이 아니라는 것을 나타내고 있다. 이것에 관련된 일반법칙은 정보의 세밀한 부분을 기억하는 것보다는 정보를 일반화 (개념화) 한 것을 기억한다는 것이다. 예를 들면 독자가 이 글을 다 읽으면 '일반화된 개념이 기억된다' 라고 하는 내용이 독자의 기억에 남는 것이고 표현의 개개의 단어는 망각될 것이다. 재생에는 기억과 반대의 과정이 작용한다. 결국 먼저 개념을 상기한 후에 언어를 취급하는 기구가 문구를 짜맞춘다. 이와 같은 점으로부터도 사람의 기억은 전자공학적인 데이터 data 기록과는 확실히 다르다.
사람이 추상적 또는 구상적 사고를 언어로 나타낼 수 있고 이와 같이 기억된다고 하는 것은 동물 (유인원도 포함) 의 기억과는 현저히 다름을 나타내고 있다. 사람에게만 가능한 언어로 부호화된 정보의 기억은 사람과 동물 양쪽에 가능한 비언어정보의 기억에 부가된 것이라고 추정된다. 이와 같이 복잡하기 때문에 동물실험의 결과를 그대로 사람의 기억과정의 해석에 적용시키는 것은 곤란하다.
정보를 기억한 기본적 기구의 이해는 현재 불충분하지만 기억은 실험적으로 구별되는 몇 개의 단계를 밟아서 일어나는 것으로 생각된다. 이와 같은 소견에 의하면 사람의 기억은 단기와 장기의 적어도 2종류의 기억으로 성립한다. 단기의 기억정보 (예: 물어보았을 뿐인 전화번호 등) 는 연습에 의해 장기기억으로 바뀌어 보이지 않으면 곧 잃어 버리고 만다. 단기기억이 한번 장기기억으로 바뀌어 보내지면 장기간 반복해서 나온다. 이렇게 해서 이루어진 기억흔적 소위 잠재적 기억상 engram 은 사용할 때마다 강화된다. 이와 같은 기억흔적의 확립은 기억된 사항을 잃어 버리기 어렵게 안정화하는 것이고 기억의 고정 consolidation 이라고 한다.
종래의 단기, 장기기억의 개념은 적어도 사람의 언어적 기억 verbal human memory 의 점에서 현재의 지식과는 맞지 않는다. <그림 16> 에 나타난 것과 같이 기억된 정보는 최초로 감각성기억 sensory memory 으로서 1 초 이내에 제것으로 된다. 감각성기억정보는 언어가 부호화된 후에 단기기억인 1 차 기억 primary memory 으로 바뀌어 보내진다. 1 차 기억에 머무르는 시간은 보통 짧게 2 ~ 3 초이지만 주의를 집중한 반복연습에 의해 시간을 연장할 수 있다. 연습에 의해 장시간 지속하는 대량의 기억인 2 차 기억 secondary memory 으로 오는 정보가 바뀌어 보내지는 것이 용이하다.
2 차 기억에 있어서 망각은 앞에 학습된 내용이나 새로운 학습의 내용에 의한 학습과정의 간섭에 의해 일어날 것이다. 앞의 학습내용에 의한 억제를 순행성 억제 proactive inhibition, 새로운 학습내용에 의한 억제를 역행성 억제 retroactive inhibition 라고 한다. 보통 앞에 학습된 내용이 망각되는 것이 많으므로 순행성 억제가 중요하다고 생각된다. 이와 같은 시야에서 보면 대부분은 이전에 배운 것이 순서대로 망각 되어지는 것이 옳게 여겨진다.
질병이나 외상에 의해 다른 기억이 상실되었을 때에는 자기의 이름이나 읽고 쓰기 외에 일상 사용하는 기능 등에 의해 외랫동안 반복 사용되었기 때문에 소실되지 않는 기억흔적이 있다 (역행성 건망증). 이들 기억흔적은 3 차 기억 tertiary memory (그림 16) 이라고 하는 특수한 형태의 기억으로서 저정되어 있다고 생각된다. 앞에서 설명한 장기기억은 이 도식에서는 1 차 기억과 3 차 기억의 양자에 대응해 있다.

<그림 16> 감각성 기억으로부터 3 차 기억까지의 정보흐름의 모식도
각 기억의 유지 지속시간과 망각의 기전을 나타낸다. 각 수준에서 유지된 자료의 일부만 다음과 같이 안정수준에 이른다. 반복 (연습) 에 의해 1 차 기억으로부터 2 차 기억으로의 바뀌어 보내지는 것이 촉진된다. 그러나 이것은 절대필요한 조건은 아니고, 또한 바뀌어 보내지는 것을 보증하는 것도 없다
새로 주어진 정보가 학습되지 않고 그 정보가 장기간 기억되지 않는 상태를 선행성 건망증 anterograde amnesia 이라고 한다. 이와 같은 환자 (종종 만성 알코올중독 환자) 는 발병 이전에 획득한 2 차 또는 3 차 기억은 거의 정상이고 1 차 기억도 바로 기능이 되고 있다. 그러나 1 차 기억으로부터 2 차 기억으로 가는 정보가 바뀌어 전달되지 않는다 (그림 16).
해마 hipocampus 및 그 외의 대뇌변연계통 limbic structure 이 양측성으로 파괴되거나 외과적으로 절제되면 완전히 동시에 불가역적인 선행성 건망증이 일어나므로 이 영역이 기억이 바뀌어 보내지는 가장 중요한 역할을 하고 있을 것이다.
뇌가 손상을 받기 이전에 기억된 내용을 상기할 수 없는 상태를 역행성 건망증 retrograde amnesia이라고 한다. 그 원인으로서는 기계적 충격 (진탕 concussion), 뇌졸중발작, 전기쇼크 (치료용 또는 사고), 마취 등이 잘 알려져 있다. 어느 것이나 뇌의 기능이 전반적으로 장애되는 것으로 특히 어느 부분 또는 어느 기능의 장애가 역행성 건망증을 불러일으키는지는 현재 불분명하다. 그러나 예를 들면 회복기에 어쩔 수 없이 망각된 것을 생각해 내는 것으로 기억의 내용을 완전히 상실해 버릴 뿐만 아니라 2 차 기억을 끌어내는 것이 장애되기 때문이라고 생각된다. 한편 3차 기억은 원칙적으로 위독한 역행성 건망증의 경우에는 영향을 받지 않는다.
학습의 신경적 기초에 관한 단순하고 동시에 구체적인 추론으로 아래와 같은 것이 있다. 정보는 최초로 공간적 및 시간적인 배열을 취해 반향흥분 reverberation excitation 의 형태로 동적 기억흔적 ynamic engram 으로서 기억된다. 그 회로의 흥분은 반향회로의 연접에 형태학적 변화를 가져오고 동적 기억흔적은 형태적 기억흔적 structural engram 으로 고정된다. 기억의 내용은 이 반향회로의 연접활성화로 상기된다.
학습의 내용을 유지하기 위해서는 이것을 반복해서 의식 속을 통과시키지 않으면 안되지만 이와 같은 주관적 체험과 흥분의 반향회로의 개념과는 잘 일치한다. 이와 같은 흥분의 반향회로의 가능성을 나타내는 형태학적 및 전기생리학적 소견도 있지만 이들 현상의 학습과정으로의 관여에 대한 상세함은 명확하지 않다.
반복자극의 중간 및 자극 후의 연접전달효율의 변화에 대해서는 앞에서 설명했다. 특히 반복자극 후 증강 posttetanic potentiation 은 예를 들면 해마 등의 흥분성 연접은 몇시간도 지속된다. 이와 같은 연접특성의 변화는 형태적 기억 흔적 형성에 따른 신경계통의 변화에 기초를 둔다고 생각된다. 이와 같은 생각은 반복자극 후 증강이 짧은 시간만 지속하지 않고 척수에서는 영속적인 학습이 인정되지 않는 사실과 일치한다. 생쥐 mouse 를 출생시부터 눈을 사용하지 못하도록 수술로 안구를 절제하든가 어두운 곳에서 사육하면 시각영역의 연접에 조직학적으로도 기능적으로도 변성이 안된다. 이 경우 사용 불충분하기 때문에 연접의 효율이 저하되는 것이고 앞에서 설명한 개념을지지한다.
학습 중의 뇌파나 뇌의 일반적 전압변화의 연구는 많지만 학습과 기억의 신경기구의 해명에 기여하는 것은 적다. 실험성적의 해석에 모순을 가져오는 것도 있고, 본 장에서는 상세히 설명하지 않는다.
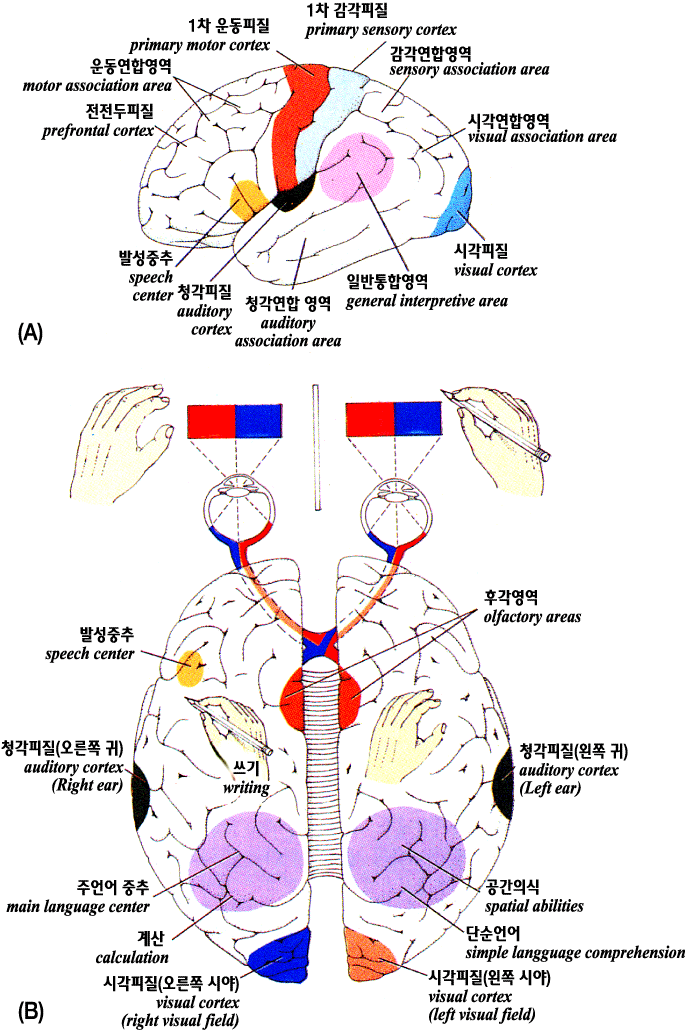
<그림 17> 고위학습 및 기억중추
A: 뇌의 외측모양은 좌반구 표면의 주요 통합 및
연합영역의 위치를 나타낸다
B: 분리증후군을 가지고 있는 대뇌반구 사이의 활동부분
설명
4) 기억흔적 engram 의 생화학적 (분자적) 기구
유전정보의 기억 genetic memory 이 핵산 (DNA) 에 부화화되어 짜넣어진다고 하는 연구나 면역학적 기억 immunological memory 의 연구 성공은 신경회로에 의한 기억 neural memory 의 기본인 분자수준의 변화를 찾아내어 연구 하려는 자극으로 되었다. 이와 같은 분자수준의 변화는 기억흔적의 기초라고 생각되고 세포내 또는 세포막의 구성성분인 단백질의 변화는 그일례일 것이다.
학습에 의해 신경세포 또는 신경교세포의 리보핵산 (RNA) 이 변화하는지 아닌지에 대해서 수많은 실험이 행해져왔다. RNA 는 세포내의 단백합성에 중요하고 그 변화는 세포단백의 구성에 변화를 가져온다. RNA 의 전체 질량 및 RNA 를 구성하는 4 개의 nucleotide 염기의 질량비를 측정하는 고등기술에 의해 어떤 종류의 학습과정에 의해 이들 염기의 질량비가 변화하는 것을 알았고, 이 변화는 단순한 비특이적 변화이지만 가능성도 있다. 한편 학습된 행동을 다른 개체로 옮겨갈 가능성을 나타내는 실험이 되어 왔다. 이것은 훈련된 동물의 일군의 뇌로부터 RNA 를 추출해서 훈련되지 않은 군에 주사하는 실험이지만 planaria 와 같은 하등동물에서도 어류나 포유류에 있어서도 이와 같은 가능성을 나타내는 확증은 발견되지 않았다.
기억의 생화학적 기초를 해명하는 실험 중에서 다음 두 가지는 설명할 가치가 있다. 한 가지는 위에서 설명한 방법과 전혀 반대로 예를 들면 RNA 합성 내지 단백합성을 actinomycin 이나 promycin 으로 억제시킴으로써 세포내 또는 세포막의 형태적 기억흔적의 형성을 장애하는 것도 있다. 이 실험은 한편에서는 성공했지만 다른 쪽에서는 단백합성의 비특이적 억제가 기억흔적 형성의 장애뿐만 아니라 세포기능 전체의 장애를 가져온다고 하는 반론을 부정할 수 없다.
다른 한 가지는 전기쇼크에 의한 벌을 주는 훈련으로 자연의 성질과는 반대로 어두운 장소를 피하게 된 흰쥐 rat 의 뇌로부터 어떤 polypeptide 를 추출해서 이것을 보통의 흰쥐나 생쥐, 금붕어에 주입시켰더니 밝은 장소에 오래 있게 된 것과 같은 실험에서 이 polypeptide 는 스코토포빈 scotophobin (그리스어의 암흑 scotos 과 공포 phobos 에서 유래함) 이라 하고 15 개의 아미노산으로 합성도 된다. 이와 같은 소견이 의미하는 것은 명확하지 않는 것이다. 어느 쪽이든지 그 결과가 12 년 전부터 알려졌는데도 관계없이 이와 같은 실험은 아직 확립된 것은 아니고 또한 행동의 학습에 관한 정보매체 information carrier 로 생각되는 새로운 대형분자도 발견되지 않았다. 더욱이 scotophobin 에 관해서는 아미노산 연쇄가 뇌하수체 호르몬의 하나인 부신피질 자극호르몬 (ACTH) 과 유사하다는 것, 또는 ACTH 는 생체의 각성이나 주의력을 증강시키는 것으로 ACTH 자체가 행동을 개선 했다고도 생각된다. 따라서 scotophobin 자체가 학습에 관여하는 물질일 가능성은 의문스럽다.
지금까지 소위 기억촉진물질로서 glutamine 산 glutamate, 항cholinestrase, choline 작동성 약품, strychine, picrotoxin, tetrasole, caffeine, RNA 등이 주장되어온 것과는 관계 없이 약리학적 방법에 의해 지금까지 지성이나 기억을 필요로 하는 작업의 달성도를 직접 동시에 특이적으로 개선할 수는 없었다. 보다 빨리 좋은 학습, 기억유지, 기억력 (회상, 상기) 을 얻었다고 하는 우리들의 여망을 이용하려는 사람도 있으므로 의심스러운 과학적 주장의 말에 대한 가짜 약에 조심할 필요가 있다.
앞에서 설명했지만 연합피질의 두정엽과 측두엽의 영역은 일부는 언어활동에 관여하고 있고 또한 일부는 외계 및 신체의 공간적, 형태적 인식에 관여한다. 이들을 기능면에서 보면 좌우의 양반구의 사이에는 흥미있는 차이가 보인다. 전두엽의 기능에 대한 자세한 설명은 현재 곤란 하지만 전두엽은 대뇌변연계통 (종특이적 행동의 제어에 관여한다) 과 밀접히 사아호연락되고 있으므로 전두엽 역할의 하나로 본능적 행동양식의 학습에 의한 제어가 고려된다. 이와 같은 생각은 전두엽의 손상을 받은 환자가 충동적으로 되거나, 비억제적, 과민, 다행 등으로 되거나 하는 등 심리적으로 불안정하게 되는 사실로부터도 뒷받침된다.
전두엽 손상 예의 임상소견으로부터 전두엽 기능을 어느 정도 관찰할 수 있다. 이와 같은 많은 예는 지능은 정상을 나타내지만 다른 한편으로는 의욕의 결여, 확고한 의지의 결여, 전망을 갖는 계획성이 결여 등의 인격변화가 종종 일어난다. 더욱이 많은 환자는 의지할 곳이 없고, 거칠고 촌스럽고, 무분별, 성실하지 모사고, 노하기 쉽고, 사회적 고민 social trouble (예: 직장 등의 고민) 에 말려들 게 된다.
이들 환자는 운동을 시키면 그 동작에 집착하는 경향이 있고 다른 동작을 하도록 명령해도 앞의 동작을 계속한다. 즉 한번 개시한 동작에 집착하여 고집하는 것을 보속증 perseveration 이라고 한다. 이와 같은 보속경향은 학습의 실험에서도 인정되고 환자는 일련의 자극 중 어떤 자극과 그에 앞선 자극의 식별곤란을 나타낸다. 위에 설명한 전두엽 손상환자의 행동은 앞에 형성된 기억흔적 engram 이 그 바로 뒤에 오는 다음 정보를 위한 장소를 점령하고 있는가 하는 인상을 준다. 즉 순행성억제가 증강되고 있다는 인상을 준다.
전두엽 손상환자는 필요성에 의해서 행동을 변화시키는 것이 곤란하다. 외부로부터의 강제에 대한 반응이 감약되고 외부로부터와 자기의 내부로부터의 동기가 대립할 때에 그 한편으로부터 다른 쪽으로 재빠르게 적절히 대처할 수 없다. 위에서 설명한 행동관찰소견은 앞에서 설명한 해부학적인 섬유연락의 소견과도 일치한다. 전두엽은 본능행동의 학습, 더욱이 억제와 외적 및 내적 동기의 조화에 관여한다. 침팬지를 처음으로 하는 포유동물의 실험도 똑같은 성적을 나타냈다. 사람 또는 동물의 소견은 행동의 계획에 전두엽이 지도적 역할을 하고 있다고 하는 가설을 시사하고 있다.
유효한 향정신약의 발견 이전에는 어떤 종류의 정신병의 치료에, 전두엽과 시상의 섬유연락을 절단하는 수술 (백질절단 leucotomy) 이 쓰였다. 이 수술은 항상 시비가 논의되었지만 오늘 날에는 이미 정당한 것으로 보지 않는다. 이 수술은 1940 년경에 최초로 행해졌고 뇌조직의 파괴나 제거에 의해 인간행동을 일생을 통해 바꾸는 어떤 정신외과의 획기적인 선구자로 되었다. 넓은 의미에서는 정신외과에는 전기쇼크요법, 장기간의 향정신약요법, 뇌내에 전극설치 등도포함된다. 그 이유는 이들이 불가역적인 뇌조직의 변화를 가져와 얻은 것이다.
뇌의 기능에 대한 현재의 지식이 너무도 적은 것을 생각하면 정신외과적 치료법의 근거는이론적으로 말하는 것보다 경험적인 것이다. 현재 가장 잘 행해지는 정신외과 수술은 대상회 (뇌량의 위에 위치해 있는 변연계통에 속해 있는) 를 절제하는 대상회절제술 cingulotomy 이고 심한 불안상태 및 우울증 등의 정신장애나 감당하루 없는 통증을 경감시키기 위해 행한다. 변연계통 핵의 하나인 편조체핵을 파괴하는 편도핵 절제술 amygdalotomy 은 공격적 행동을 억제하기 위한 최후의 수단으로 사용된다. 단지 이 수술도 인격에 본질적인 변화를 불가역적으로 더하는 것으로 중대한 의심이 더해진다.
이와 같은 수술의 경우에도 다른 모든 외과 기술 진보의 경우와 똑같이 적정한 규범을 작성해서 새로운 지식을 인류의 이익을 위해서만 사용되도록 보증하는 것은 사회의 책임이다.